I. — Introduction
Le gisement de diatomite dit « de Murat » (Cantal), situé autour du point de coordonnées Lambert III : x = 642,80 ; y = 3314,00 (longitude 02°52'50''E, latitude 45°07'30''N), est connu depuis 1875 et exploité en grand depuis 1901. Il constitue une ellipse de 0,8x1,3 km, d'une épaisseur moyenne de 20 mètres ; il est recouvert de dépôts alluviaux ou périglaciaires et surtout d'une énorme accumulation de moraine de fond atteignant 25 mètres de puissance. Les exploitations ont par ailleurs rencontré trois intrusions basaltiques, dont l'une datée à 5,4 Ma, qui perturbent le fond du gisement et thermométamorphisent les diatomites. Le gisement de diatomite est donc daté du Messinien, et noté m3D sur la carte géologique (Durand et Rey, 1964 ; Goër de Hervé et Tempier, 1988).
Les espèces fossiles de la macroflore livrée par ce gisement particulièrement riche ont été précédemment mentionnées et/ou décrites par Marty in Piton et Théobald (1935), Privé-Gill (1987), Roiron (1991) et Legrand (2003).
De nouvelles récoltes de macroflore effectuées dans la diatomite de Murat nous amènent aujourd'hui à compléter l'inventaire précédent (Legrand, 2003).
II. — Matériel et méthodes
Au cours de la période 2002-2009, nous avons poursuivi les récoltes de macroflore dans la carrière exploitée par la société CELITE, dans la continuité de notre précédent inventaire (Legrand, 2003). Par ailleurs, nous avons récolté des échantillons sur les tas de diatomite entreposés par la société CECA, qui exploite une carrière située sur le même gisement, mais à plusieurs centaines de mètres à l'Est-Sud-Est de la précédente.
Au cours de la période 2004-2009, le front de taille de la carrière exploitée par la société CELITE s'est déplacé vers l'Est-Sud-Est, en direction de la chapelle Sainte-Reine. A cette occasion, le niveau de base de la formation de diatomite, qui constituait le fond (plancher) de la carrière, a été mis en exploitation ; bien que déjà connu au moins par sondages, ce niveau n'avait jamais été exploité industriellement auparavant. Nous avons donc procédé à l'inventaire de la macroflore de ce niveau (exploité surtout au cours des premières années, puis stocké en tas dans la carrière) que nous nommerons dans la suite du texte « niveau de base à Synedra ». Ce niveau ne devrait d'ailleurs plus être exploité, car trop riche en argiles.
Dans cette partie exploitée du gisement, les couches de diatomites sont réparties de la façon suivante, de la base vers le sommet :
- niveau de base à Synedra : riche en argiles, contenant principalement des diatomées Synedra, quelques diatomées Cyclotella, et quelques autres espèces de diatomées, très riche en macroflore (notamment en feuilles d'Alnus stenophylla Saporta & Marion), et riche en insectes fossiles ;
- niveau à diatomées Synedra principalement, très fortement varvé, très pauvre en macroflore ;
- niveau à diatomées Melosira principalement, plus ou moins riche en macroflore ;
- niveau terminal à diatomées Cyclotella principalement, plus ou moins riche en macroflore, livrant aussi des insectes.
Ces niveaux à diatomées ne peuvent être appréciés de manière sure qu'au microscope, le faciès de terrain pouvant être identique d'un niveau à l'autre. Ainsi, ces niveaux sont matérialisés par l'exploitant sur le terrain au fur et à mesure de l'exploitation. Il faut aussi noter que cette stratigraphie est celle de la carrière CELITE exploitée au cours de cette période, mais que cette stratigraphie n'est pas retrouvée sur l'ensemble du gisement de diatomite. Ainsi, le niveau à Cyclotella s'est rétréci au fur et à mesure de l'avancement de l'exploitation CELITE ; il disparaît totalement dans la carrière exploitée par CECA où ne subsiste principalement que le niveau à Synedra (A. Ferrand, société CELITE, comm. pers.) ; ceci est peut-être dû à un rabotage par les glaciers, l'ensemble du gisement de diatomite étant recouvert par une moraine de fond.
Dans cet article, nous mettrons l'accent sur deux points :
- la description de formes fossiles et d'espèces qui n'avaient pas été signalées jusqu'à présent, et qui ont été récoltées dans l'ensemble du gisement de diatomite de Murat ;
- l'inventaire de la macroflore du niveau de base à Synedra ; pour les autres niveaux, les formes fossiles de macroflore récoltées mais déjà connues ne seront pas traitées de nouveau, sauf précisions particulières apportées par les nouvelles récoltes.
Les méthodes relatives à l'étude des formes fossiles et à leur détermination sont identiques à celles utilisées pour l'inventaire initial (Legrand, 2003). Nous rappelons que :
- les espèces fossiles sont nommées telles que décrites dans la littérature, et les espèces actuelles sont suivies par convention du qualificatif fossilis lorsque la forme fossile rencontrée semble s'identifier à une espèce vivante actuelle ;
- la nomenclature botanique retenue pour les unités de rang supérieur au genre est celle préconisée par Judd et al. (2002).
III. — Nouvelles formes fossiles récoltées
Nous décrivons ci-dessous les nouvelles formes fossiles récoltées dans la diatomite de Murat, et apportons des précisions sur quelques formes précédemment décrites.
TRACHEOPHYTES A SPORES (PTERIDOPHYTES)
Fougères leptosporangiées
POLYPODIACEAE s. l.
Pteridium aquilinum (Linné) Kuhn (= Pteris aquilina Linné) fossilis
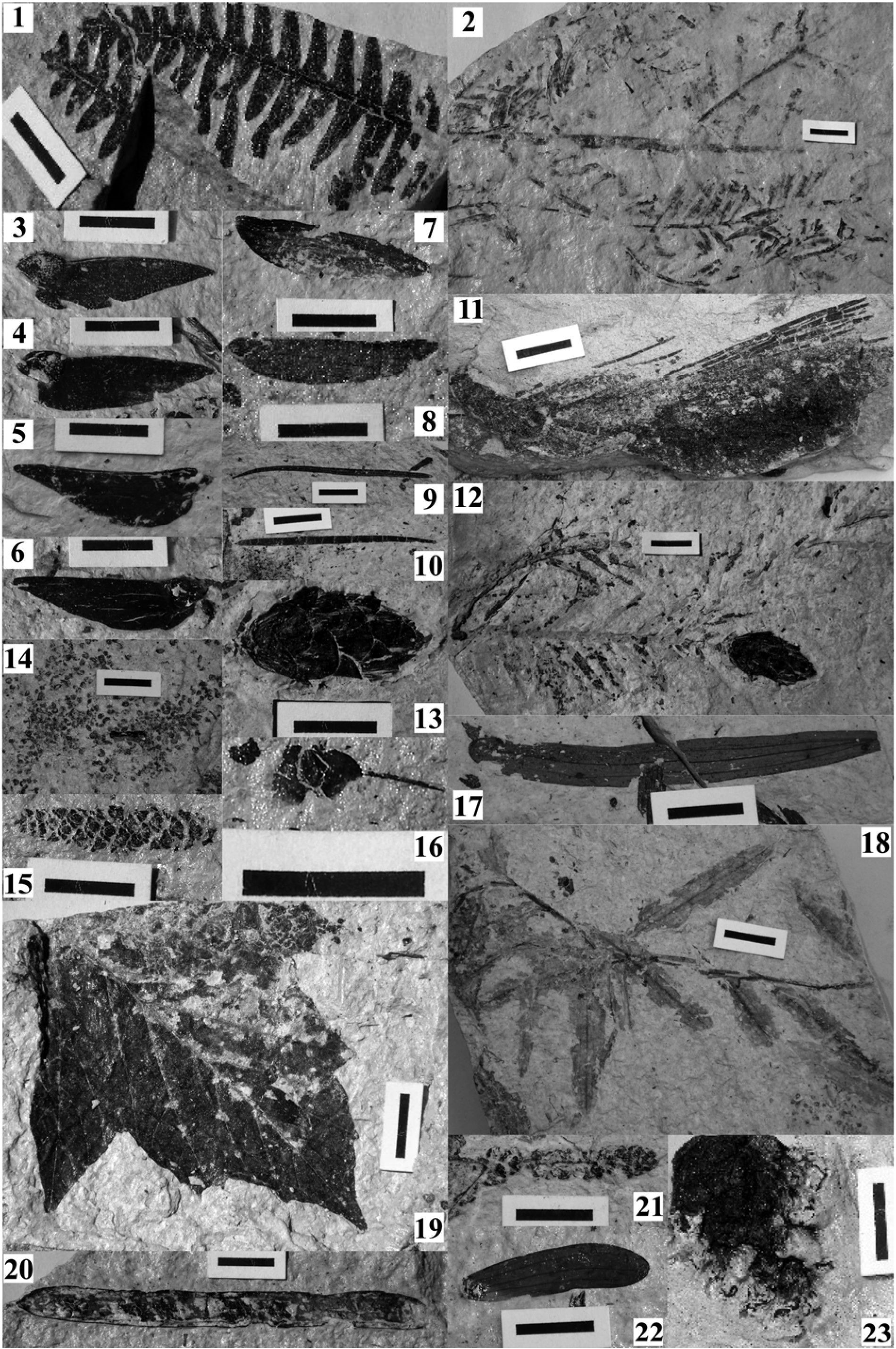
(Pl. I, fig. 1)
Un petit échantillon comportant deux pennes de dernier ordre incomplètes a été récolté dans le niveau à Melosira. Les pinnules mesurent jusqu'à 1 cm de long, pour environ 3,5 mm de large à la base. Cet échantillon est rapporté à la fougère aigle actuelle.
Laurent et Marty (1927) avaient déjà signalé cette espèce dans le gisement de Cheylade (Cantal), qui est en partie contemporain de celui de Murat (Roiron, 1991).
SPERMATOPHYTES NON ANGIOSPERMES (« GYMNOSPERMES »)
Conifères
PINACEAE
Piceoxylon sp. et/ou Pinuxylon sp.
Une douzaine de bois fossilisés en lignite a été récoltée dans la diatomite de Murat ; ces bois ont fait l'objet d'une étude en lames minces par C. Privé-Gill (Université Pierre-et- Marie-Curie, Paris). Ces bois sont en général de conservation très médiocre. Le spécimen PL-MUR11, et sans doute le spécimen PL-MUR13, appartiennent au genre Piceoxylon Gothan ou au genre Pinuxylon Gothan, bois fossiles de Pinaceae.
Le genre Piceoxylon désigne les bois fossiles voisins des actuels Picea, Larix et Pseudotsuga. Ce sont les deux premiers genres surtout qui sont délicats à distinguer par l'anatomie du bois ; le troisième est assez différent car muni d'épaississements spiralés sur les parois des trachéides. Le genre Pinuxylon est utilisé pour les bois fossiles rattachés aux seuls Pinus actuels. Les deux genres fossiles se distinguent par l'épaisseur des parois des cellules sécrétrices des canaux sécréteurs : parois épaisses chez Piceoxylon, minces chez Pinuxylon. Dans les bois de Murat, on peut voir des canaux sécréteurs horizontaux, mais les canaux verticaux sont assez obscurs, et il est impossible de décider si les parois des cellules sécrétrices sont épaisses ou minces. On ne peut non plus distinguer les détails nécessaires tels que les diverses ponctuations des trachéides, ni la présence et donc l'aspect des trachéides horizontales des rayons. Il est donc impossible de trancher entre Piceoxylon et Pinuxylon, et d'aboutir à une détermination spécifique (Privé-Gill, comm. pers.).
Pour ce qui concerne des bois fossiles rapportés à ces deux genres :
- Piceoxylon mosquense (Kraus) Schönfeld, voisin de l'actuel Picea abies (Linné) Karsten, a été identifié dans le Miocène supérieur de La-Bastide-du-Fau (Cantal) et dans le Pliocène volcano-sédimentaire des Monts-Dore, vers La- Tour-d'Auvergne (Puy-de-Dôme) (Privé-Gill, 1987) ;
- Pinuxylon parryoides (Gothan) Kräusel emend. Van der Burgh, voisin de l'actuel Pinus pinea Linné, a été déterminé dans divers gisements du Miocène supérieur du Cantal (Privé-Gill, 1987).
Abies sp. et/ou Cedrus sp.
Des ailes de graine, dépourvues de leur graine et isolées, ont été rencontrées notamment dans le niveau de base à Synedra. Elles sont à rapprocher des genres Abies et Cedrus qui sont connus dans le gisement par d'autres macrorestes.
Abies ramesi Saporta (Pl. I, fig. 2)
Des aiguilles isolées d'Abies ramesi ont été précédemment signalées dans la diatomite de Murat (Roiron, 1991 ; Legrand, 2003).
Nous avons aussi récolté dans les déblais de la carrière un rameau feuillé, malheureusement de mauvaise conservation : rameau principal de 11 cm de long, comportant deux séries de rameaux opposés, le plus long atteignant une dizaine de centimètres (ramification verticillée dans un plan). Les aiguilles mesurent 17-19 mm de longueur, avec coussinet d'insertion sur le rameau bien visible.
Cedrus sp. et/ou Picea sp.
Des aiguilles isolées, courtes, aciculaires et à section quadrangulaire, sont rencontrées de manière dispersée dans la diatomite de Murat. Ces aiguilles sont à rapprocher des genres Cedrus et Picea qui sont connus dans le gisement par d'autres macrorestes.
Pinus sp. 2
(Pl. I, fig. 7 et 8)
Nous avons précédemment décrit et figuré une graine de Pinus sp. (Legrand, 2003 ; planche V, figure 12) ; nous avons depuis récolté de nombreuses graines identiques en taille, dont de nombreuses graines microspermes, qui mesurent un total d'environ 1,5 cm de longueur de la pointe de la graine à l'extrémité de l'aile. Nous nommerons désormais ces graines « Pinus sp. 1 ».
Par ailleurs, nous avons récolté trois graines microspermes du même genre botanique, mesurant au total 2,5-2,8 cm de longueur, avec une aile de 5-6 mm de large ; ces graines beaucoup plus grandes appartiennent très certainement à une espèce distincte, et nous les nommerons « Pinus sp. 2 ». L'une de ces graines provient du niveau de base à Synedra, les deux autres des déblais de la carrière.
Par comparaison avec les espèces actuelles, on pourrait rapprocher les graines de Pinus sp. 1 de Pinus sylvestris Linné, et les graines de Pinus sp. 2 du groupe du Pinus nigra Arnold.
Pinus sp
(Pl. I, fig. 23)
Un cône femelle de mauvaise conservation, mesurant 3,5 cm de hauteur pour 2,5 cm de largeur a été récolté dans les déblais de la carrière.
Pinus sp.
(Pl. I, fig. 11)
Un rameau feuillé, de mauvaise conservation, a été récolté dans le niveau de base à Synedra. Il correspond probablement à l'extrémité d'un rameau en cours d'élongation, et porte des aiguilles longues d'au moins 4,3 cm ; en raison du piètre état de conservation, il est impossible de préciser le nombre d'aiguilles par faisceau.
Pinus sp.
(Pl. I, fig. 9 et 10)
Deux aiguilles isolées ont été récoltées dans le niveau de base à Synedra, l'une de 4,4 cm de long, l'autre de 5,5 cm.
Pseudolarix schmidtgenii Kräusel
Synonyme : Pseudolarix amabilis (J. Nelson) Rehder fossilis
(Pl. I, fig. 3, 4, 5 et 6)
Une douzaine de graines ailées à la forme typique et identique à l'espèce actuelle ont été récoltées principalement dans le niveau de base à Synedra, quelques-unes se trouvant dans les déblais de la carrière. Longueur totale (de la pointe de la graine à l'extrémité de l'aile) 2,7-2,8 cm ; largeur de l'aile 7-8 mm, la loge de la graine faisant environ 5x3,5 mm.
Florschütz (1925) a été le premier à reconnaître des macrorestes fossiles pliocènes de ce genre en Europe, qu'il a rapportés à la seule espèce actuelle qui vit en Chine : Pseudolarix amabilis (J. Nelson) Rehder (anciennement P. kaempferi [Ldl.] Gordon). Ensuite, Hickel (1932) a décrit des écailles de cône et une graine ailée dans le Pliocène d'Alsace (France), qu'il a aussi rapprochées de l'espèce actuelle. Enfin, Kräusel (1938) a décrit des graines ailées miocènes comme holotype de Pseudolarix schmidtgenii, d'une morphologie tout à fait identique à l'espèce actuelle.
A la suite, plusieurs auteurs ont signalé d'autres macrorestes (graines ailées et écailles de cônes) dans le Mio- Pliocène d'Europe, qui ne diffèrent pas morphologiquement de l'espèce actuelle. Ces macrorestes ont été rapportés à l'espèce fossile de Kräusel et/ou à l'espèce actuelle ; on peut citer notamment : Mädler (1939), Martinetto (2001 ; avec, géologiquement parlant, les derniers macrorestes fossiles datés du Pliocène terminal au Pléistocène inférieur, avant la disparition de ce genre botanique d'Europe), Kunzmann et Mai (2005), Kvacek et Teodoridis (2007).
La distribution des fossiles de Pseudolarix dans l'hémisphère Nord au cours des temps géologiques a été compilée par Manchester et al. (2009) et surtout par LePage et Basinger (1995). Ces derniers indiquent que, parmi tous les fossiles du genre en Amérique du Nord et Eurasie (et qui ont reçu divers noms selon leur provenance géographique ou géologique), seules deux espèces peuvent être distinguées sur la morphologie de la bractée de l'écaille du cône : une espèce secondaire représentant une lignée éteinte d'ampleur limitée P. wehrii Gooch, et une espèce principale très stable morphologiquement tout au long de son évolution et tout à fait similaire à l'espèce actuelle P. amabilis ; ils déclarent ainsi que rien ne justifie de distinguer ces restes fossiles comme une espèce différente de l'espèce actuelle.
Les pollens de Pseudolarix peuvent être confondus avec ceux du genre Pinus (Ferguson, 1967), et nous ne retiendrons donc comme mention en France que celle de Zanni et Ravazzi (2007), basée sur de nouvelles investigations permettant de les différencier ; ces auteurs établissent ainsi la présence de pollens de Pseudolarix dans des sédiments lacustres âgés d'environ 3 Ma à La Gratade (massif des Monts Dore, Massif Central).
En dehors des pollens, le genre Pseudolarix n'a, à notre connaissance, été signalé en France que dans le Pliocène d'Alsace, sous la forme d'écailles de cônes et de graines ailées (Hickel, 1932 ; Geissert, 2005) ; les graines récoltées à Murat sont donc les plus anciens macrorestes connus en France.
Tsuga europaea Menzel
(Pl. I, fig. 12 et 13)
Un rameau feuillé, comportant trois ramifications apparemment non verticillées, et un cône en position terminale d'un rameau, a été récolté dans le niveau à Melosira. Seul le cône est bien conservé, une grande partie de la matière organique des axes et feuilles ayant malheureusement disparu de l'empreinte et de la contre- empreinte. Les aiguilles sont aplaties avec nervure centrale bien marquée, de 11-18 mm de long sur 2-2,5 mm de large, à extrémité apparemment arrondie sur les empreintes ; leur insertion sur le rameau est très peu visible mais semble se faire par un court pédicelle. Le cône femelle mesure 2,4 cm de long sur 1,2 cm de large ; il est inséré à l'extrémité d'un rameau court d'environ 12 mm ; les écailles du cône sont arrondies et les bractées non visibles. Ce fossile (détermination L. Kunzmann) est rapporté à Tsuga europaea Menzel.
Les espèces actuelles du genre Tsuga habitent l'Amérique du Nord et l'Asie (Debazac, 1964 ; Gaussen, 1966), mais ce genre est connu en Europe depuis le Crétacé supérieur jusqu'au Pléistocène (Ferguson, 1967). En dehors des pollens, différentes formes fossiles de Tsuga ont été décrites en Europe, notamment Tsuga europaea Menzel qui est un cône du Mio-Pliocène (Gaussen, 1966, 1967 ; Kunzmann et Mai, 2005) ; des feuilles ont aussi été rattachées à cette espèce (Mädler, 1939).
Le cône récolté dans la diatomite de Murat est d'une taille assez proche de celle de l'holotype et d'autres spécimens rapportés à cette espèce (Kunzmann et Mai, 2005), bien qu'il s'agisse très probablement d'un cône immature puisqu'attaché à son rameau, les cônes de Tsuga étant caducs à maturité (Kunzmann, comm. pers.).
A noter que des pollens de Tsuga avaient déjà été signalés dans la diatomite de Murat (Durand et Rey, 1964).
ANGIOSPERMES
Graines indéterminées
(Pl. I, fig. 14)
Des graines isolées ou en amas, de forme arrondie ou légèrement ovale, et de 1 à 1,5 mm de diamètre sont très nombreuses dans le niveau de base à Synedra. Il est possible que la forme fossile que nous avions précédemment rapportée avec doute à une inflorescence de Vitis sp. ? (Legrand, 2003 ; planche X, figure 7) soit en fait à rapprocher de ces graines indéterminées ; dans les deux cas, la fossilisation ne permet pas une détermination plus précise.
Monocotylées
ALISMATALES POTAMOGETONACEAE
Potamogeton crispus Linné fossilis
(Pl. I, fig. 18 et Pl. II, fig. 3)
Nous avions précédemment décrit une feuille entière ovale oblongue rapportée au genre Potamogeton (Legrand, 2003, p. 30 et pl. V fig. 14). Nous avons trouvé d'autres spécimens fossiles différents que nous rapportons à l'actuel Potamogeton crispus L., à feuilles toutes submergées, largement répandu sur le globe (Coste, 1937). Ces tiges feuillées ont été récoltées dans le niveau de base à Synedra, et sont souvent de mauvaise conservation.
Tiges rameuses comprimées larges de 1 à 2 mm. Feuilles sessiles, longueur 3-4,5 cm, largeur 7-8 mm ; nervures parallèles longitudinales : une nervure principale centrale souvent bien marquée, et deux nervures de chaque côté de la nervure principale ; entre ces nervures parallèles, présence de nervures secondaires formant des échelons obliques entre les nervures parallèles, bien visibles sur l'un des spécimens récoltés.
Teodoridis (2007) a passé en revue un certain nombre de formes fossiles de Potamogeton, qui sont d'une manière générale assez rares dans le Tertiaire. Laurent et Marty (1923) avaient par ailleurs souligné qu'il est délicat d'aboutir à une spécification des feuilles fossiles de Potamogeton.
« Complexe des magnoliidées »
LAURALES
LAURACEAE
Sassafras ferrettianum Massalongo
(Pl. II, fig. 1)
Nous avons précédemment figuré une feuille entière provenant de la diatomite de Murat (Legrand, 2003 ; planche IX, figure 11).
Nous avons depuis trouvé deux feuilles trilobées et tout à fait caractéristiques de l'espèce, l'une dans le niveau à Cyclotella, l'autre dans les déblais de la carrière.
Tricolpées (dicotylées vraies)
PROTEALES et autres « Tricolpées primitives »
PLATANACEAE
Platanus aceroides Goeppert
(Pl. I, fig. 19)
Une feuille incomplète (la base manque) récoltée dans le niveau de base à Synedra ; les 3 lobes supérieurs sont conservés, au moins en partie. Nervation principale visible, des nervures latérales aboutissant au niveau de grosses dents peu nombreuses et peu prononcées du bord du limbe. En l'absence d'éléments particulièrement discriminants, mais en raison de la présence de grosses dents sur le bord du limbe du lobe terminal (le seul bien conservé), nous préférons attribuer cette feuille de Murat aux platanes avec doute plutôt qu'aux érables (genre Acer).
Platanus aceroides est une forme fossile polymorphe qui regroupe les caractères de divers platanes actuels (Laurent, 1908 ; Depape, 1922) et signalée par Laurent (1908) dans le gisement de Niac (Cantal), daté du Tortonien et donc à peine plus ancien que Murat (Roiron, 1991). Pour les espèces actuelles, il faut d'ailleurs remarquer l'extraordinaire polymorphie foliaire, ainsi que la présence d'hybrides, ce qui entraîne des difficultés importantes voire l'impossibilité d'une détermination spécifique sur les seuls caractères foliaires (Jovet et Kerguélen, 1990).
SAXIFRAGALES
HALORAGIDACEAE
Myriophyllum sp
(Pl. II, fig. 9 et 10)
Tiges feuillées immergées récoltées dans le niveau de base à Synedra, avec des feuilles atteignant au moins 2,5 cm de longueur.
Forme fossile proche des différentes espèces actuelles du genre Myriophyllum, déjà signalé dans la diatomite de Murat par Marty (in Piton et Théobald, 1935), mais que nous n'avions pas retrouvé précédemment.
SANTALALES
VISCACEAE
Viscophyllum pliocaenicum (Engelhardt) Mädler
(Pl. I, fig. 17 et 22)
Feuilles entières linéaires-rubanées, de 2,6-6 cm de long et 6-8 mm de large, à sommet arrondi et base tronquée, comportant 3-5 nervures principales plus ou moins parallèles bien visibles, avec nervures secondaires peu visibles.
Mädler (1939), après étude des cuticules, a attribué aux Viscaceae, sous le nom Viscophyllum pliocaenicum, des feuilles de 1,4-4,6 cm de long et 1,5-4 mm de large, précédemment rapportées au genre Potamogeton (Potamogetonaceae). En France, cette espèce a été identifiée dans le Pliocène d'Alsace, sous la forme de feuilles de 2,13,5 cm de long et 5-8 mm de large (Kvacek et al., 2008). Bien que les feuilles de Murat soient d'une dimension supérieure à celles précédemment décrites, et en l'absence d'une étude des cuticules permettant de confirmer cette attribution, nous les rapportons à l'espèce fossile Viscophyllum pliocaenicum.
MALPIGHIALES
SALICACEAE
Populus nigra Linné fossilis
(Pl. I, fig. 21)
Deux chatons lâches associés à une grande feuille repliée en deux (hauteur du limbe supérieure à 12 cm ; demi-largeur de 5 cm) ont été récoltés dans le niveau de base à Synedra. Ces chatons mesurent environ 2,5 cm de longueur, plus un pédoncule d'au moins 1 cm.
FABALES
FABACEAE (= LEGUMINOSAE)
Fabaceae : fruit
(Pl. I, fig. 20)
Une gousse de Légumineuse a été récoltée dans le niveau à Cyclotella. Elle mesure 7 mm de largeur pour 8 cm de longueur (une petite partie d'une extrémité manque) ; elle contient 5 ou 6 graines visibles par transparence, ovales, d'une dimension de 5-7x2-4 mm (étant donné la fossilisation, il est difficile de dire si la plus grosse graine est unique ou si deux graines sont superposées).
Pour mémoire, des feuilles de Cercis sp. aff. Cercis siliquastrum Linné fossilis étaient les seuls restes de Légumineuses signalés jusqu'à présent dans la diatomite de Murat (Legrand, 2003).
ROSALES
ROSACEAE
cf. Photinia sp.
(Pl. II, fig. 4)
Une feuille récoltée dans le niveau de base à Synedra présente un pétiole de 1,7 cm de long, et un limbe de 5,5 cm de hauteur, 2,1 cm de largeur, en coin à la base, acuminé au sommet, bordé de petites dents dirigées vers le haut, surtout dans la moitié supérieure du limbe.
Nous rapportons cette feuille au genre Photinia avec réserves, genre déjà signalé par Roiron (1991) dans la diatomite de Murat. Il faut en effet noter que les feuilles des Rosacées présentent à la fois des caractères communs aux différents genres et une grande variabilité dans le genre ; ceci rend difficile la comparaison et l'attribution des feuilles fossiles à un genre vivant ou à une forme fossile déjà décrite (Laurent, 1904-1905).
Prunus pereger Unger
(Pl. II, fig. 2)
Une feuille au pétiole incomplet, avec un limbe de 8,3 cm de haut pour 5 cm environ de large, récoltée dans le niveau de base à Synedra, est conforme au spécimen figuré et décrit par Laurent (1904-1905), provenant des cinérites du Pas-de-la- Mougudo (Cantal), gisement en partie contemporain de celui de Murat (Roiron, 1991).
Cette forme fossile est proche de différentes espèces actuelles de Prunus d'Amérique, d'Europe et d'Asie ; la feuille est plus particulièrement proche de celle de l'actuel merisier Prunus avium L., mais ne possède pas les deux glandes situées sur le pétiole à la base du limbe (Laurent, 1904-1905).
FAGALES
BETULACEAE
Alnus sp.
(Pl. I, fig. 15)
Chatons mâles récoltés dans le niveau de base à Synedra, d'une longueur de 1,7-2,5 cm, pour une largeur de 4-5 mm.
Betula sp.
(Pl. I, fig. 16)
Graines ailées de petite taille (environ 4 ou 5 mm de diamètre, aile comprise), récoltées dans le niveau de base à Synedra.
SAPINDALES
SAPINDACEAE (y compris ACERACEAE)
Acer pseudoplatanus Linné fossilis
(Pl. II, fig. 6 et 7)
Une dizaine de feuilles typiques à cinq lobes dentés, proches de l'érable sycomore actuel, récoltées dans le niveau de base à Synedra ; hauteur du limbe 8-12 cm, largeur 10,515 cm ; pétiole pouvant atteindre une longueur supérieure ou égale à 8 cm.
Acer tricuspidatum Bronn = Acer trilobatum Al. Braun
(Pl. II, fig. 5)
Une feuille incomplète (la base manque) a été récoltée dans le niveau de base à Synedra. Elle est constituée de 3 lobes séparés par des sinus aigus. Malgré la base manquante, nous rapprochons cette feuille de l'Acer tricuspidatum, déjà signalé à Murat par Roiron (1991). Comme indiqué par Brice (1965), le bord du limbe est finement denté (petites dents inégales, aiguës, dirigées vers l'avant sur notre spécimen). Cette espèce est aussi donnée comme polymorphe (Brice, 1965 ; Roiron, 1991).
CORNALES
CORNACEAE
Cornus sanguinea Linné fossilis
(Pl. II, fig. 8)
Une feuille incomplète (le haut manque) mais suffisamment typique a été récoltée dans le niveau de base à Synedra ; sa demi-largeur est de 2 cm, et la longueur du limbe est estimée à 7 cm ; 4 paires de nervures secondaires sont visibles : les trois premières s'insèrent sur la nervure médiane dans la moitié inférieure du limbe, et la dernière paire, très écartée des précédentes, s'insère dans la moitié supérieure.
Cette feuille est rapportée au cornouiller sanguin actuel. Chez le cornouiller mâle (Cornus mas Linné), toutes les nervures secondaires sont situées dans la partie inférieure de la feuille, avec un espacement régulier entre paires, et manquent totalement dans la moitié supérieure (Laurent, 1908). L'espèce fossile Cornus distans N. Boulay possède un nombre de nervures secondaires plus important (Grangeon, 1956).
AQUIFOLIALES
AQUIFOLIACEAE
Ilex geissertii Kvacek, Teodoridis & Wang
Roiron (1991) avait rapproché des feuilles de houx de la diatomite de Murat de l'actuelle espèce chinoise Ilex cornuta Lindl. & Paxt., tandis que d'autres feuilles semblaient se rapprocher aussi de l'actuelle espèce européenne Ilex aquifolium L. (Legrand, 2003).
Par ailleurs, la forme fossile Ilex boulei Saporta avait été décrite par Laurent (1904-1905, 1908), dans le Mio-Pliocène du Cantal, comme une variété d'I. aquifolium portant un champignon parasite observé sur des Ilex chinois. Legrand (2003) avait donc suggéré d'utiliser la dénomination Ilex boulei pour les houx fossiles mio-pliocènes voisins d'I. aquifolium mais pouvant aussi recouvrir d'autres espèces euro-asiatiques.
Tout récemment, Kvacek et al. (2009) ont décrit une nouvelle forme fossile, Ilex geissertii, à partir de matériel fossile mio-pliocène précédemment attribué à I. cornuta et à I. aquifolium. Ils ont de plus figuré et rattaché à cette espèce des feuilles provenant de la diatomite de Murat.
Nous substituons donc cette nouvelle espèce Ilex geissertii aux anciennes dénominations (Ilex boulei et Ilex sp. aff. I. cornuta).
APIALES
APIACEAE (= UMBELLIFERAE ; y compris ARALIACEAE)
Hedera helix Linné fossilis
Nous avons signalé précédemment dans la diatomite de Murat la présence d'une feuille de lierre, entière, à la morphologie des feuilles de rameau florifère de lumière (Legrand, 2003). Nous avons retrouvé plusieurs feuilles identiques dans le gisement de diatomite, ainsi que deux feuilles arrondies et une feuille triangulaire, toutes présentant des amorces de lobes, dans le niveau de base à Synedra.
IV. — Mise à jour de l'inventaire de la macroflore
Dans le tableau I, nous donnons :
- d'une part, l'inventaire des formes fossiles rencontrées dans le niveau de base à Synedra, avec le nombre d'échantillons récoltés pour chaque forme fossile ; ce nombre est uniquement indicatif ; en effet, de nombreux fragments plus ou moins identifiables n'ont pas été récoltés lorsqu'il s'agissait de formes fossiles déjà connues ;
- d'autre part, la liste complète des formes fossiles rencontrées jusqu'à présent dans le gisement de diatomite de Murat, avec la référence bibliographique des mentions (citation seule des espèces ou citation avec illustration).
Si l'on compare l'inventaire du niveau de base à Synedra avec l'inventaire précédent (Legrand, 2003), on constate que certains taxons sont absents du niveau de base, d'autres du niveau supérieur. Il est difficile de dire s'il s'agit d'une véritable absence ou d'un manque de fossilisation, les récoltes effectuées (et soumises à l'avancement d'une carrière en exploitation) ayant montré que la présence de végétaux n'est pas homogène au sein d'un même niveau. Par ailleurs, en paléontologie « l'absence de preuve ne vaut pas preuve d'absence », et il apparaît par conséquent hasardeux, avec les données actuelles, de rechercher une évolution de la macroflore pendant la durée de formation du gisement (estimée à 50000 ans par Fournier, 1965) et liée par exemple à des modifications environnementales. En effet, une telle discussion devrait impérativement prendre en compte la paléopalynologie. Or les données palynologiques restent très fragmentaires sur le gisement de Murat (bibliographie in Legrand, 2003), et aucune étude niveau par niveau n'y a été menée ; ces données seront de plus maintenant très difficiles à établir en raison de l'avancement de l'exploitation du gisement et de la disparition de certains niveaux.
V. — Conclusions
Le « niveau de base à Synedra » qui constitue la base du gisement de diatomite apparaît très riche à la fois en nombre de fossiles qu'il renferme et en nombre d'espèces, puisqu'il a livré 67 formes fossiles différentes, représentant un total de 52 espèces : 6 Gymnospermes, 2 Angiospermes Monocotylédones et 44 Angiospermes Dicotylédones.
La description d'une vingtaine de nouvelles formes fossiles récoltées dans l'ensemble du gisement de diatomite de Murat nous permet d'ajouter 10 nouvelles espèces à la flore du gisement : 1 Ptéridophyte, 3 Gymnospermes, 1 Angiosperme Monocotylédone et 5 Angiospermes Dicotylédones. Parmi celles-ci, les graines de Pseudolarix schmidtgenii Kraüsel, espèce fossile similaire à l'actuel Pseudolarix amabilis (J. Nelson) Rehder de Chine, apparaissent comme un élément remarquable.
L'inventaire de la macroflore fossile de la diatomite de Murat est donc maintenant au total de 83 espèces : 3 Ptéridophytes, 11 Gymnospermes (dont 1 bois fossile), 4 Angiospermes Monocotylédones (en comptabilisant deux espèces fossiles de Bambusa) et 65 Angiospermes Dicotylédones (dont 1 bois fossile, et en comptabilisant deux espèces de Carpinus, l'espèce de forme C. suborientalis Saporta recouvrant les deux espèces C. orientalis Miller fossilis et C. betulus Linné fossilis). Avec l'abondance des spécimens récoltés, ceci en fait sans nul doute l'un des plus riches gisements en végétaux fossiles miocènes du Massif Central.
Remerciements. — Nous remercions tout particulièrement Mme Catherine Privé-Gill (Paléobiodiversité et Paléoenvironnements, Université Pierre-et-Marie-Curie, Paris) pour l'étude des bois fossiles, M. Lutz Kunzmann (Senckenberg Naturhistorische Sammlungen Dresden, Museum für Mineralogie und Geologie, Dresden, Allemagne) et M. Aljos Farjon (Royal Botanic Gardens, Kew, Royaume-Uni) pour les échanges d'informations sur les conifères fossiles et les déterminations de Pseudolarix et Tsuga, ainsi que M. James F. Basinger (University of Saskatchewan, Saskatoon, Canada) et M. David K. Ferguson (Institut für Paläontologie, Université de Vienne, Autriche) pour leur aide bibliographique. Nous remercions enfin Melle Denise Brice (Société géologique du Nord) et M. Paul Roiron (Institut de Botanique, Université de Montpellier) pour la relecture et l'amélioration du manuscrit.
Nous remercions vivement les sociétés CELITE et CECA qui nous ont accordé les autorisations de prospection et de récolte sur le gisement en exploitation et plus particulièrement M. Yves Aufauvre et M. Pierre Bila, directeurs successifs de CELITE, M. Alain Ferrand, responsable de l'exploitation CELITE, qui nous a toujours accueilli très cordialement et fait partager sa connaissance du gisement, et M. Henri David, directeur de CECA.
Nous remercions aussi Mme Nathalie Peynon-Legrand pour son aide lors des récoltes.
Tableau I
| Formes fossiles récoltées | Diatomite de Murat Références bibliographiques |
Niveau de base à Synedra Nombre de fossiles récoltés |
| Ptéridophytes | ||
| Asplenium viride Huds. fossilis (penne avec pinnules) | 4 | |
| Equisetum sp. aff. E. fluviatile L. fossilis (tige) | 4 | |
| Pteridium aquilinum (L.) Kuhn fossilis (penne avec pinnules) | 5 | |
| Gymnospermes | ||
| Abies sp. ? ou Cedrus sp. ? (base de cône) | 4 | |
| Abies sp. et/ou Cedrus sp. (aile de graine) | 5 | 8 |
| Abies ramesi Saporta (graine, aile de graine) | 3 | |
| Abies ramesi Saporta (aiguille isolée) | 3, 4 | |
| Abies ramesi Saporta (rameau feuillé) | 5 | |
| Cedrus sp. et/ou Picea sp. (aiguilles) | 4, 5 | + |
| Cedroxylon sp. Kraus (bois) | 2 | |
| Cedrus miocenica Lauby (graine ailée) | 4 | 1 |
| Cedrus miocenica Lauby (écaille de cône femelle) | 4 | |
| Cedrus miocenica Lauby (chaton mâle) | 4 | 2 |
| Cedrus miocenica Lauby (rameau défeuillé) | 4 | |
| Glyptostrobus europaeus (Brongniart) Heer (rameau feuillé) | 3 | |
| Juniperus sp. section sabina (rameau) | 4 | 4 |
| Picea sp. (graine ailée) | 3, 4 | 15 |
| Picea sp. (cône femelle) | 3, 4 | 1 |
| Picea sp. (aiguille) | 3 | |
| Piceoxylon sp. et/ou Pinuxylon sp. (bois) | 5 | |
| Pinus sp. 1 aff. P. sylvestris L. fossilis (graine ailée) | 3, 4, 5 | 23 |
| Pinus sp. 2 aff. P. nigra Arn. fossilis (graine ailée) | 5 | 1 |
| Pinus sp. (cône femelle) | 5 | |
| Pinus sp. (rameau feuillé) | 5 | 1 |
| Pinus sp. (aiguille isolée) | 5 | 2 |
| Pseudolarix schmidtgenii Kräusel (= P. amabilis [J. Nelson] Redher fossilis) (graine ailée) | 5 | 9 |
| Sequoia langsdorfii (Brongniart) Heer (rameau feuillé) | 3 | |
| Tsuga europaea Menzel (rameau feuillé avec cône femelle) | 5 | |
| Angiospermes | ||
| Graines indéterminées, isolées ou en amas | 5 | 15 |
| Angiospermes Monocotylédones | ||
| Monocotylédones (feuilles) | 3, 4 | nb |
| Bambusa sp. (feuille) | 3 | |
| Bambusa lugdunensis Saporta & Marion (feuille) | 4 | 7 |
| Potamogeton sp. (feuille ovale-oblongue) | 4 | |
| Potamogeton crispus L. fossilis (tige feuillée submergée) | 5 | 5 |
| Angiospermes Dicotylédones | ||
| Acer sp. (samare) | 3, 4 | 3 |
| Acer decipiens Al. Braun (feuille) | 4 | 3 |
| Acer laetum pliocenicum Saporta & Marion = Acer integerrimum (Viv.) Mass. (feuille) | 1, 3, 4 | 15 |
| Acer opalus Mill. (= Acer opulifolium Vill.) fossilis (feuille) | 3 | |
| Acer palmatum Thunberg fossilis = Acer sanctae-crucis Stur (feuille) | 3, 4 | 5 |
| Acer platanoides L. fossilis (feuille) | 3, 4 | |
| Acer pseudocampestre Unger = Acer campestre L. fossilis (feuille) | 3, 4 | |
| Acer pseudoplatanus L. fossilis | 5 | 10 |
| Acer tricuspidatum Bronn (= Acer trilobatum Al. Br.) (feuille) | 3, 5 | 1 |
| Alnus sp. (strobile femelle) | 3, 4 | 13 |
| Alnus sp. (chaton mâle) | 5 | 4 |
| Alnus glutinosa (L.) Gaertner fossilis (feuille) | 3, 4 | |
| Alnus kefersteinii Unger (feuille) | 3, 4 | 2+1? |
| Alnus stenophylla Saporta & Marion = Alnus hoernesi Stur (feuille) | 1, 3, 4 | nb |
| Alnus viridis D.C. fossilis (feuille) | 3 | |
| Berberis sp. cf. B. regeliana Koehne fossilis (feuille) | 3,4 | 2 |
| Betula sp. (écaille de chaton femelle) | 3 | |
| Betula sp. (graine ailée) | 5 | 8 |
| Betula macrophylla (Goeppert) Heer (feuille) | 3, 4 | 8 |
| Betula subpubescens Goeppert (feuille) | 4 | 7 |
| Buettneria tiliaefolia (Al. Braun) Depape (feuille) | 4 | |
| Carpinus betulus L. fossilis (fruit avec involucre) | 3, 4 | 1 |
| Carpinus orientalis Mill. fossilis (fruit avec involucre) | 3, 4 | 3 |
| Carpinus suborientalis Saporta (feuille) | 3, 4 | 7 |
| Carya minor Saporta & Marion (feuille ; foliole) | 3, 4 | 1 |
| Cedrela sp. (graine) | 4 | |
| Cedrela sp. (feuille) | 3 | |
| Celtis australis L. fossilis (feuille) | 3, 4 | 2 |
| Ceratophyllum demersum L. fossilis (tige feuillée) | 3, 4 | 1 |
| Cercis aff. Cercis siliquastrum L. fossilis (feuille) | 4 | 1 |
| Cornus sanguinea L. fossilis (feuille) | 5 | 1 |
| Corylus avellana L. fossilis (feuille) | 4 | 8 |
| Crataegus sp. série Crus-Galli et Punctata (feuille) | 4 | 1 |
| Crataegus sp. cf. C. douglasii Lindl. fossilis (feuille) | 3 | 1 ? |
| Dombeyopsis lobata Unger (feuille) | 3, 4 | |
| Fabaceae (fruit : gousse de Légumineuse) | 5 | |
| Fagus pliocenica Saporta (feuille) | 4 | |
| Fraxinus sp. (feuille) | 4 | |
| Hedera helix L. fossilis (feuille) | 3, 4, 5 | 8 |
| Ilex geissertii Kvacek, Teodoridis & Wang (feuille ; rameau avec feuilles et fruits ) | 3, 4, 5 | 2 |
| Juglans regia L. fossilis (foliole) | 1, 3, 4 | 13 |
| Laurus azorica (Seub.) J. Franco fossilis (feuille) | 4 | 2+2 ? |
| Myrica sp. (feuille) | 4 | 1 |
| Myriophyllum sp. (tiges feuillées) | 1, 5 | 10 |
| Persea indica Spr. fossilis (feuille) | 4 | 1 ? |
| Phellodendron sp. cf. P. amurense Rupr. fossilis (feuille) | 3 | |
| Cf. Photinia sp. (feuille) | 3, 5 | 1 |
| Platanus aceroides Goeppert (feuille) | 5 | 1 |
| Populus alba L. fossilis (feuille) | 4 | |
| Populus x canescens (Ait.) Sm. fossilis (feuille) | 4 | 1 |
| Populus nigra L. fossilis (feuille) | 4, 5 | 1 |
| Populus nigra L. fossilis (chaton) | 5 | 2 |
| Populus tremula L. fossilis (feuille) | 3, 4 | 7 |
| Prunus acuminata Al. Br. (feuille) | 3 | |
| Prunus microdonta N. Boulay (feuille) | 4 | 3+2 ? |
| Prunus pereger Unger (feuille) | 5 | 1 |
| Pterocarya denticulata Heer (foliole) | 4 | |
| Quercoxylon böckhianum (Felix) Müller-Stoll & Mädel (bois) | 2 | |
| Quercus hispanica Rérolle (feuille) | 1, 3, 4 | 2 |
| Quercus kubinyi (Kov.) Berger (= Castanea kubinyi Kov.) (feuille) | 3 | |
| Quercus sp. cf. Quercus macranthera Fisch. & Mey. fossilis (feuille) | 3, 4 | |
| Rosa sp. cf. Rosa californica Cham. & Schlechtd. fossilis (foliole) | 3, 4 | |
| Salix sp. à feuilles étroites (feuille) | 4 | 1 |
| Sapindus falcifolius Al. Braun (foliole) | 4 | 1 |
| Sassafras ferrettianum Massalongo (feuille entière, feuille trilobée) | 4, 5 | |
| Sorbus aria (L.) Crantz fossilis (feuille) | 4 | 3 |
| Tilia sp. (bractée florale) | 3, 4 | |
| Tilia tomentosa Moench. fossilis (feuille) | 3, 4 | |
| Ulmus sp. (samare) | 3, 4 | 1 |
| Ulmus braunii Heer (feuille) | 1, 3, 4 | 10 |
| Ulmus sp. cf. Ulmus fulva Michx. fossilis (feuille) | 3, 4 | 2 |
| Viscophyllum pliocaenicum (Engelh.) Mädler | 5 | 2 |
| Vitis sp. ? (inflorescence) | 4 | |
| Vitis praevinifera Sap. (feuille) | 4 | |
| Vitis vivariensis N. Boulay aff. Vitis vinifera L. fossilis (feuille) | 4 | 1 |
| Zelkova ungeri Kovats aff. Z. carpinifolia (Pall.) K. Koch fossilis (feuille) | 1, 3, 4 | 1 |
| Zelkova ungeri Kovats aff. Z. serrata (Thunb.) Mak. fossilis (feuille) | 3, 4 | 1 |
Inventaire de la macroflore de la diatomite de Murat
Première colonne : inventaire des formes fossiles rencontrées dans le gisement ; Deuxième colonne : références bibliographiques des espèces citées 1= Marty in Piton et Théobald, 1935 ; 2= Privé-Gill, 1987 ; 3= Roiron, 1991 ; 4= Legrand, 2003 ; 5= Legrand, cet article ; Troisième colonne : inventaire des formes fossiles du niveau de base à Synedra, avec le nombre de spécimens récoltés ; + = présence ; nb = nombreux échantillons
Inventory of the fossil flora from the diatom beds at Murat
First column : inventory of the fossil forms in this site ; Second column : bibliographical references of the cited species : 1= Marty in Piton et Théobald, 1935 ; 2= Privé-Gill, 1987 ; 3= Roiron, 1991 ; 4= Legrand, 2003 ; 5= Legrand, (this paper) ; Third column : inventory of the fossil forms collected in the Synedra diatom bed located at the basis of the geological level, with the number of specimens collected (+ = present ; nb = a lot of specimens)